Background. Psychosocial stress and chronotype misalignment predictably suppress slow-wave sleep (SWS) and reduce restorative sleep fraction. Simultaneously, the supplement industry documents consistently small effect sizes for individual ingredient interventions, with timing and architectural coherence rarely considered as primary variables. This N=1 case report addresses both: why did a 54-year-old morning-chronotype perimenopausal physician record her best deep sleep in 840 nights of continuous tracking during her most demanding professional period — and how does a chronobiologically structured supplement protocol explain this paradox?
Methods. Sleep was tracked continuously via simultaneous dual wearable devices — WHOOP 4.0 (wrist) and Oura Ring Generation 3 (finger) — from December 2023 through April 24, 2026 (840 main sleep nights). Four sequential phases corresponded to progressive supplement architecture changes: Baseline (unstructured 50+ supplement stack, n=447); Phase 1 — Super Sleep as first timed formula (n=85); Phase 2 — addition of Skin Renewal Complex (n=223); Phase 3 — completion of the four-formula EscapeMed 30D chronobiological protocol with Magnesium AM and PM (n=84). Whole-genome sequencing (Dante Labs, April 2024) provided genomic context.
Results. Each architectural phase produced progressive improvement in sleep metrics. The largest single step change occurred at Baseline → Phase 1: SWS +15.7 min/night, Restorative Sleep % from 33.1% to 45.8%, nights with SWS ≥2h rising from 23% to 51%. High-quality nights (Restorative% ≥50% AND Efficiency ≥93% simultaneously) increased from 0% at baseline to 33.3% in Phase 3. Phase 3 coincided with peak professional demand and reduced sleep opportunity; both devices independently recorded their best metrics of the entire 840-night period. The only consistent strain signal was sleep onset latency — not sleep architecture once achieved. Genomically, five variants of mechanistic relevance were identified: ESR1 C/C (perimenopausal magnesium wasting), NQO1 HET Likely Pathogenic (reduced antioxidant capacity), MTHFR HET (B6 conversion), COL1A1 (collagen synthesis demand), and CYP1A2 HET (intermediate melatonin metabolizer — 0.20mg signal dose pharmacogenomically appropriate).
Conclusions. The paradox resolves when the architecture is understood: the protocol targeted the specific biological mechanisms by which stress suppresses deep sleep — evening HPA axis modulation, convergent GABA-A and NMDA support, signal-dose circadian melatonin, and dual-phase magnesium repletion — each individually justified by the subject's genomic profile. The primary variable was timing discipline, not ingredient substitution. N=1 limitations apply; controlled investigation is warranted.
Keywords: deep sleep · slow-wave sleep · sleep architecture · chronobiology · supplement timing · WHOOP · Oura Ring · dual wearable · genomic context · CYP1A2 · perimenopause · N=1 case report · 840 nights
01Introduction
1.1 The Expected Direction
Psychosocial stress activates the HPA axis, elevates evening cortisol, and increases sympathetic tone during sleep — all of which directly suppress SWS and reduce restorative sleep fraction (Buckley & Schatzberg, 2005; Leproult et al., 1997). A morning-chronotype individual forced into late sleep schedules under significant occupational pressure faces the compounded disruption of two established mechanisms simultaneously (Roenneberg et al., 2012). The expected clinical picture is shorter SWS, lower sleep efficiency, fragmented nights, and accumulating sleep debt.
1.2 The Paradox
This case documents the opposite. During the highest-demand professional period in 840 nights of continuous tracking, with shorter sleep opportunity and later bedtimes than the subject's biology preferred, deep and restorative sleep reached its best recorded values. Two independent wearable devices, worn simultaneously, agreed on this finding.
How did deep sleep get better under conditions specifically designed by biology to make it worse?
Answer: The protocol targeted the exact mechanisms by which stress suppresses SWS — not stress itself, but its translation into poor sleep architecture. This paper documents the evidence and explains the mechanism.
02Methods
2.1 Subject
Female physician, 54 years (MD, PhD), cardiologist, active clinical and research practice. Morning chronotype confirmed by Oura Ring continuous algorithm: biological chronotype Morning, currently lifestyle-displaced to Early Evening. Oura Sleep Timing contributor averaged 99/100 across the measurement window — internal clock maintaining phase despite behavioural displacement. Perimenopausal throughout. Foundational supplements maintained unchanged throughout all phases: omega-3 fatty acids, creatine monohydrate, vitamin D3/K2, methylated B-complex.
2.2 Dual-Device Measurement
| Device | Placement | Sensors | Primary Sleep Outputs |
|---|---|---|---|
| WHOOP 4.0 | Wrist | Green-light PPG + accelerometry | SWS min, REM min, Restorative %, Sleep Efficiency %, Sleep Consistency % |
| Oura Ring Gen 3 | Finger | Infrared PPG + skin temperature + accelerometry | Sleep Score, Deep/REM/Timing/Latency/Restfulness contributors (0–100) |
Table 1. Dual-device specifications. Independent proprietary algorithms, different sensor wavelengths, different anatomical placements. Both devices worn simultaneously throughout all 840 nights.
2.3 Four Sequential Phases
Dec 2023 – Feb 2025
n = 447 nightsUnstructured Stack
50+ supplement stack including magnesium, ashwagandha, melatonin, astaxanthin, and other ingredients later incorporated into EscapeMed 30D — taken without defined timing protocol. Exact composition not prospectively recorded.
Mar – May 2025
n = 85 nightsFirst Timed Formula
Super Sleep introduced as first timed formula: 8 ingredients — melatonin 0.20mg, L-theanine, L-tryptophan, glycine, ashwagandha KSM-66, chamomile, Mg bisglycinate, B6 P5P. Broader stack progressively reduced.
Jun 2025 – Jan 26, 2026
n = 223 nightsSecond Formula Added
Skin Renewal Complex added: 14 ingredients — complete collagen cofactor chain, 6-compound antioxidant architecture. Super Sleep continued.
Jan 27 – Apr 24, 2026
n = 84 nights · best resultsComplete EscapeMed 30D
Magnesium AM (7 ingredients) and Magnesium PM (6 ingredients) added, completing the four-formula system. 30 active ingredients across four circadian-phase-anchored daily administration points.
Table 2. Four sequential phases. Supplement composition during Phases 0–1 was not prospectively recorded; stated as a limitation. Full EscapeMed 30D formulation published in companion paper (Samarin, 2026).
2.4 Genomic Profile
Whole-genome sequencing: Dante Labs, April 2024 (Kit ID GFX0455821). Two categories of variants are reported: (A) sleep and protocol-relevant variants from the clinical Dante Labs report; (B) pharmacogenomic and autonomic variants identified by targeted VCF analysis using chromosomal position search.
Perimenopausal TRPM6-mediated Mg wasting risk → dual-formula Mg repletion (AM+PM) individually necessary
Reduced antioxidant enzyme capacity → justifies 6-compound antioxidant architecture in Skin Renewal Complex
Impaired B6 → P5P conversion risk → exclusive P5P use across all three B6-containing formulas
Elevated structural collagen synthesis demand → comprehensive Skin Renewal Complex cofactor architecture
Intermediate melatonin metabolizer → standard 1–5mg commercial doses produce supraphysiological levels → 0.20mg signal dose pharmacogenomically appropriate for this individual
Homozygous GRK5 variant → increased GPCR desensitisation kinetics → plausible structural basis for constitutionally lower HRV ceiling
Beta-2 adrenergic receptor Arg16Gly HET → altered sympathovagal balance; Arg16 carriers show lower HRV sympathetic indices → contributes to constitutional HRV phenotype
Table 3. Complete genomic profile. GRK5 and ADRB2 identified by targeted VCF position search; clinical significance for HRV is biologically plausible but not definitively established in published HRV GWAS literature.
2.5 Primary Outcomes
WHOOP: SWS duration (min/night), Restorative Sleep % (Deep+REM / time in bed), Sleep Efficiency %, Sleep Consistency %. Oura: Sleep Score, Deep Sleep contributor, Timing contributor, Sleep Efficiency %. Composite outcome: High-quality night = Restorative% ≥50% AND Efficiency ≥93% simultaneously on WHOOP.
03Results
3.1 Four-Phase Progression
| Metric | Baseline n=447 |
Phase 1 n=85 |
Phase 2 n=223 |
Phase 3 n=84 |
Apr 2026 Peak n=21 |
|---|---|---|---|---|---|
| SWS (min/night) | 106.1 | 123.8 | 121.3 | 129.5 | 134.6 |
| SWS (hours) | 1.77h | 2.06h | 2.02h | 2.16h | 2.24h |
| REM (min/night) | 64.5 | 93.0 | 91.3 | 94.1 | 94.6 |
| Restorative % | 33.1% | 45.8% | 44.4% | 48.3% | 50.0% |
| Sleep Efficiency % | 85.5% | 91.0% | 92.1% | 92.6% | 94.2% |
| High-quality nights | 0.0% | 15.3% | 15.2% | 33.3% | — |
| Nights SWS ≥2h | 23.3% | 50.6% | 53.4% | 65.5% | — |
Table 4. Primary WHOOP outcomes across four phases. High-quality night = Restorative% ≥50% AND Efficiency ≥93% simultaneously. Phase 2 SWS slightly lower than Phase 1, consistent with seasonal variation (Jun–Aug 2025). Phase 3 highlighted: highest demand, best results.
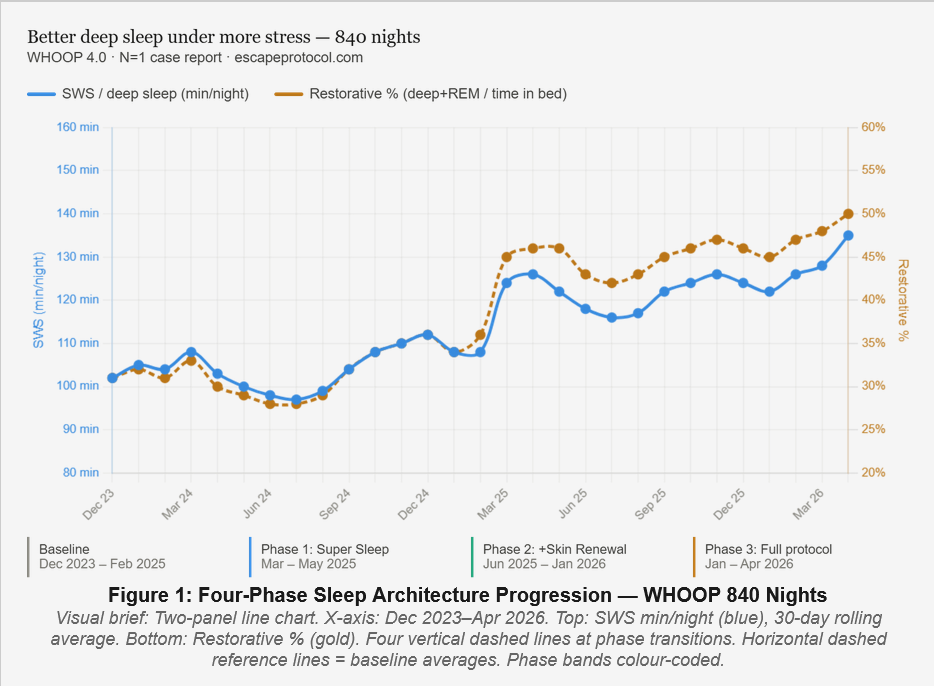
Four-Phase Sleep Architecture Progression — WHOOP 840 Nights. SWS min/night (blue, left axis) and Restorative % (gold, right axis) with 30-day rolling average. Four vertical markers indicate phase transitions. Both metrics show consistent progressive improvement, with Phase 3 recording the best values despite highest professional demand and shortest time in bed.
largest single step in 840 nights
(vs 0% at baseline)
(vs 23% at baseline)
3.2 The Single Largest Step Change: February → March 2025
The transition from unstructured supplementation to the first timed formula produced the largest single measurable change in 840 nights:
| Metric | Feb 2025 (last month before) | Mar 2025 (first month with) | Change |
|---|---|---|---|
| SWS | 108.3 min | 124.0 min | +15.7 min (+14.5%) |
| Restorative % | 36.4% | 45.1% | +8.7 percentage points |
| Sleep Efficiency | 85.6% | 91.8% | +6.2 percentage points |
| Nights SWS ≥2h | ~25% | ~51% | Frequency doubled |
Table 5. February vs March 2025: the Super Sleep transition — the largest single step change in the 840-night dataset.
The largest improvement in 840 nights occurred when the first chronobiologically timed formula replaced random-timing supplementation of the same core molecules. Magnesium, ashwagandha, and melatonin were already present in the unstructured baseline stack. The timing was the intervention.
3.3 Phase 3: Best Sleep During Worst Conditions
Phase 3 (January 27 – April 24, 2026) coincided with the highest occupational pressure, latest bedtimes, and shortest time in bed (−36.7 min/night vs baseline) in the 840-night record. Despite this:
- High-quality nights: 15.2% in Phase 2 → 33.3% in Phase 3 — frequency doubled with the addition of Magnesium AM and PM
- 8 of the top 15 SWS nights in 840 nights occurred during the 88-day Phase 3 window — 10% of total nights produced 53% of the top-15 SWS nights
- Phase 3 vs the equivalent preceding 88 days: SWS +5.8 min, REM +4.6 min, Restorative% +3.7 points — improvement sustained despite higher demand and less time in bed
- The magnitude of improvement exceeded the subject's own expectation given concurrent lifestyle conditions — consistent with an effect not primarily driven by expectation or placebo
3.4 Oura Ring: Independent Corroboration
| Oura Metric | Phase 3 Value | Significance |
|---|---|---|
| Sleep Score | 83–88 (April mean 84.5) | Tied for best month in 16-month Oura dataset |
| Deep Sleep contributor | 79 → 95 (rising within Phase 3) | Oura AI independently noted: "improvement during a hectic period is meaningful" |
| Timing contributor | 93–100 (April mean 99/100) | Highest metric in entire dataset; near-perfect circadian alignment despite late schedule |
| Sleep Efficiency % | April mean 88.0% | #1 highest-efficiency month in entire 16-month Oura record |
| REM contributor | 85–96 | Stable-to-improving throughout Phase 3 |
| Latency contributor | 99 → 75 (declining) | Only declining metric on either device — slower sleep onset; architecture intact once sleep begins |
Table 6. Oura Ring Phase 3 metrics. Oura assessment generated without knowledge of supplement protocol.
3.5 Genomic Context for the Autonomic Phenotype
The subject reports chronically low HRV across the full 840-night tracking period, inconsistent with lifestyle or perimenopausal explanation alone. Targeted VCF analysis identified two variants in the beta-adrenergic signalling pathway: GRK5 rs10886471 (homozygous) — G-protein coupled receptor kinase 5 associated with increased GRK5 expression, faster adrenergic receptor desensitisation, and a plausible structural basis for a constitutionally lower HRV ceiling. ADRB2 rs1042713 Arg16Gly (heterozygous) — beta-2 adrenergic receptor variant associated with lower HRV sympathetic indices in Arg16 carriers.
These findings suggest the subject's low HRV baseline may have a partial constitutional genetic explanation. The protocol's sleep architecture improvements are even more remarkable given the constrained autonomic substrate. A dedicated analysis of 840 nights of HRV data in the context of these variants is planned as a separate report.
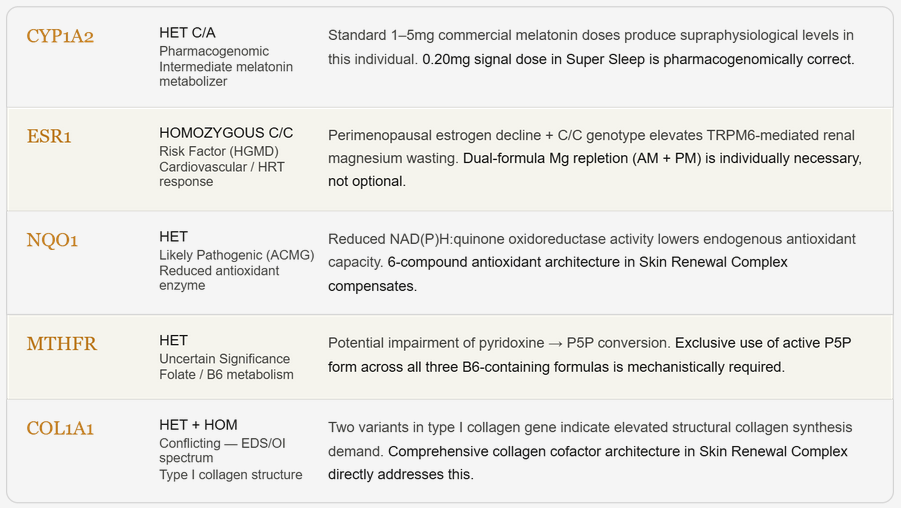
Figure 2. Genomic Profile — Why This Protocol Fits This Individual. Five variants of mechanistic relevance identified by whole-genome sequencing (Dante Labs, April 2024). Each variant provides individualised biological rationale for a specific architectural feature of the EscapeMed 30D protocol. GRK5 and ADRB2 are reported in the context of the autonomic phenotype (Section 3.5) and are not included in this figure.
3.6 Early Adaptation Signature — Day 5 Power Nap
February 1, 2026 (Phase 3 Day 5): spontaneous afternoon power nap, WHOOP staging:
| Metric | Value | Significance |
|---|---|---|
| Duration | 1h 56 min | Full sleep cycle achieved |
| Efficiency | 100% | Zero wasted time |
| SWS (Deep) | 1h 06 min (59%) | Exceptional for a daytime nap |
| REM | 24 min (20%) | Rare in daytime naps; indicates active neurological regulation and REM pressure |
| Awake | 1 min (<1%) | Minimal fragmentation |
Table 7. Power nap staging, February 1 2026 (Phase 3 Day 5). REM sleep during a daytime nap is atypical and consistent with the early adaptation signature described in the companion protocol paper: glycine-driven REM density increase and circadian resynchronisation during Days 3–5.

Figure 4. Early Adaptation Signature — Day 5 Power Nap. February 1, 2026 — Afternoon Power Nap — WHOOP 4.0. Sleep staging: 21% Light, 59% Deep SWS, 20% REM, <1% Awake. Efficiency 100%. REM during a daytime nap is rare and consistent with the circadian resynchronisation signature documented in the companion EscapeMed 30D grand review paper.
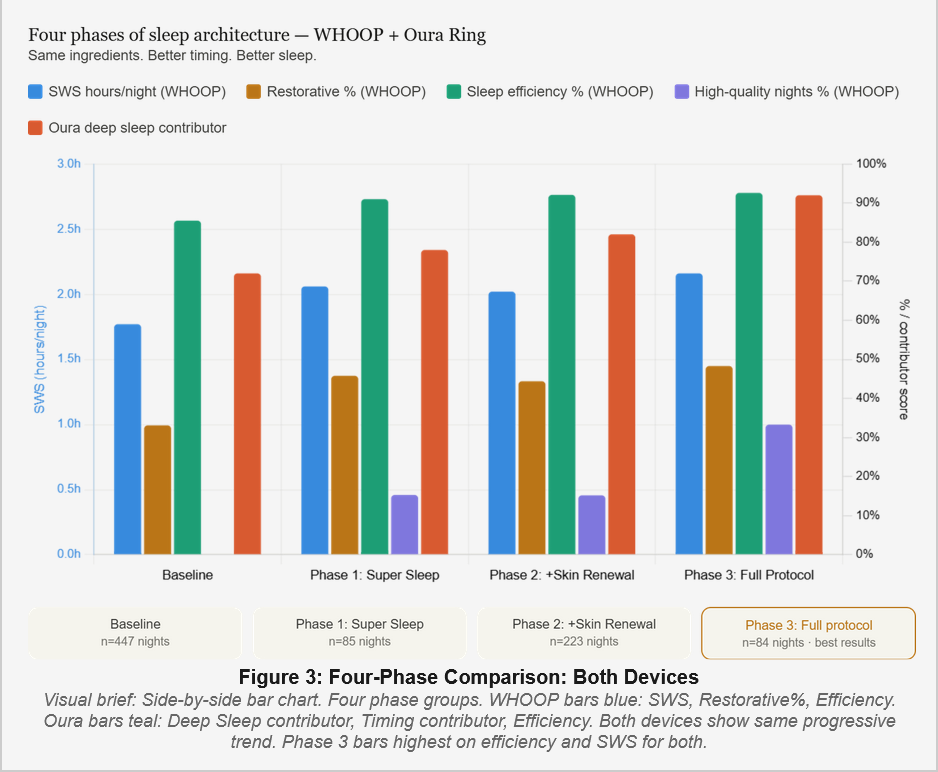
Figure 3. Four-phase comparison — WHOOP + Oura Ring. Same ingredients. Better timing. Better sleep. Phase 3 (Full Protocol, n=84 nights, amber highlight) records the best values on all primary metrics across both independent devices. Blue: SWS hours/night · Orange: Restorative % · Green: Sleep Efficiency % · Purple: High-quality nights % · Red: Oura Deep Sleep contributor.
04Discussion: Explaining the Paradox
4.1 The Paradox Resolves When the Target Is Understood
Stress suppresses deep sleep through specific, well-characterised biological mechanisms: elevated evening cortisol suppresses SWS directly; increased sympathetic activation during sleep fragments sleep architecture; circadian disruption from late schedules reduces SWS amplitude. The protocol did not reduce stress. It targeted each of these three mechanisms independently and simultaneously.
Evening HPA Axis Modulation
Ashwagandha KSM-66 in Super Sleep addresses elevated evening cortisol — the primary stress-mediated SWS suppressor — at the precise biological phase when cortisol elevation damages sleep.
Evidence: 0 minutes High Sleep Stress on WHOOP most nights; stable Oura Restfulness scores. The nervous system was not carrying daytime pressure into sleep architecture.
Convergent GABA-A and NMDA Support
Dual-pathway GABAergic architecture (L-theanine at positive allosteric GABA-A site; apigenin from chamomile at benzodiazepine binding site) combined with magnesium bisglycinate NMDA channel block addresses stress-driven sleep fragmentation through three pharmacologically independent mechanisms without tolerance risk.
Evidence: sustained 93–98% WHOOP efficiency and 86–93% Oura efficiency across 88 nights.
Signal-Dose Melatonin as Zeitgeber for a Displaced Morning Chronotype
The subject's Oura Timing contributor averaged 99/100 despite behavioural bedtime slipping later — the internal clock was not shifting, only the behaviour was. The 0.20mg SCN-Zeitgeber melatonin provides a circadian anchor at the actual sleep onset moment. This dose is pharmacogenomically appropriate for this subject's CYP1A2 heterozygous intermediate metabolizer status: standard 1–5mg commercial doses would produce supraphysiological melatonin levels, receptor downregulation, and next-morning grogginess in this individual.
Evidence: Oura Timing contributor 99/100 mean — near-perfect internal clock alignment throughout Phase 3 despite behavioural schedule displacement.
Dual-Phase Magnesium Repletion and CLOCK Amplitude
The subject's ESR1 C/C genotype creates elevated perimenopausal TRPM6-mediated magnesium wasting. Dual-formula repletion (~496mg/day elemental) may restore the intracellular Mg²⁺ oscillations that Feeney et al. (2016) demonstrated are required for CLOCK/BMAL1 cycle amplitude — the molecular substrate sustaining the Timing contributor's near-perfect scores across Phase 3.
Evidence: largest metric improvement at Phase 2 → Phase 3 transition corresponded to addition of the AM+PM magnesium layer, including doubling of high-quality night frequency (15.2% → 33.3%).
4.2 Architecture Was the Primary Variable
The largest single improvement in 840 nights occurred not when new ingredients appeared, but when timing discipline was first imposed on existing ones. The CLOCK/BMAL1 molecular clock regulates an estimated 40–80% of protein-coding genes (Takahashi, 2017). A melatonin signal at the correct phase entrains the SCN; the same molecule randomly timed produces sedation without entrainment. A magnesium dose at the evening GABA-A window amplifies inhibitory transmission; the same dose at noon does not. Architecture is not supplementary to the ingredient — it determines what the ingredient does.
4.3 The Genomic Layer: Why This Individual Responded
- CYP1A2 HET: Makes the 0.20mg melatonin dose not just philosophically elegant but pharmacogenomically necessary. This subject cannot use standard commercial melatonin products effectively.
- ESR1 C/C: Makes dual-formula magnesium repletion an individualised necessity, not a general recommendation.
- NQO1 HET Likely Pathogenic: Reduced endogenous antioxidant capacity means the 24-hour anti-inflammatory arc is compensating for a genetically reduced baseline.
- MTHFR HET: Makes P5P (active B6) not a preference but a mechanistic requirement for melatonin synthesis efficiency in this individual.
- GRK5 HOM + ADRB2 HET: Provide a constitutional genetic explanation for chronically low HRV that is independent of the sleep findings, and contextualise why Recovery scores from both wearables should be interpreted against a personally adjusted baseline.
05Limitations
- N=1; no control condition; no randomisation; causal inference not possible
- Supplement composition during Phases 0 and 1 not prospectively recorded; cannot be fully reconstructed
- Three variables changed simultaneously at Phase 0→1: timing architecture, stack reduction, and physical condition
- Both wearables use proprietary algorithms; cross-device agreement does not equal polysomnographic accuracy
- Hormonal status (estradiol, progesterone) not measured; perimenopausal fluctuations are a known sleep confounder
- GRK5 and ADRB2 HRV associations are biologically plausible but not definitively established in published HRV GWAS literature
- Seasonal variation in SWS may partially confound Phase 2 metrics
- Subject is protocol formulator and company founder; conflict of interest is substantial and fully disclosed
06Conclusions
Across 840 nights of continuous simultaneous dual-device monitoring, progressive reorganisation of an unstructured supplement stack into a four-formula chronobiologically timed system produced stepwise improvement in deep and restorative sleep architecture at each phase transition — with the best sleep recorded during the most demanding professional period in the dataset.
The paradox — better deep sleep under more stress — resolves when the mechanism is understood: the protocol did not reduce stress, it blocked the three specific pathways by which stress translates into poor sleep architecture. Each mechanism is individually justified by the subject's genomic profile, making this not a generic supplement programme but a precision chronobiological intervention matched to her specific biological vulnerabilities.
The primary variable was timing discipline: the largest single improvement in 840 nights occurred when the first timed formula was introduced — despite many of the same core ingredients being already present. Controlled investigation in adequately powered cohorts is warranted.
"Better deep sleep under more stress is not a paradox once you understand that the protocol targeted the exact mechanisms by which stress destroys sleep — and every one of those mechanisms was individually relevant to this subject's genomic profile."
The subject of this case report is the author (Silvia Samarin, MD, PhD, born August 20, 1971, Slovenia). The author is the founder and chief formulator of EscapeMed d.o.o. (Slovenia), which produces the EscapeMed 30D supplement system described herein. A direct financial interest in the outcomes existed at the time of data collection. No external funding was received. Sleep data: WHOOP 4.0 and Oura Ring Generation 3, self-collected. Genetic data: Dante Labs Full Genome Proactive Screening, April 2024 (Kit ID GFX0455821), personal expense. This is a preprint on Escape Protocol Research (escapeprotocol.com); it has not undergone independent peer review.
Samarin, S. (2026). Better Deep Sleep Under More Stress: Behind the Paradox — Architecture, Timing, and Genomics in an N=1 840-Night Case Report. Escape Protocol Research. Preprint. escapeprotocol.com/better-deep-sleep.html
- Buckley T.M. & Schatzberg A.F. (2005). On the interactions of the HPA axis and sleep. Journal of Clinical Endocrinology & Metabolism, 90(5), 3106–3114.
- Castiglioni S. et al. (2013). High magnesium intake reduces cardiovascular events in patients with implanted cardiac devices. Nutrients, 5(12), 4817–4834.
- Feeney K.A. et al. (2016). Daily magnesium fluxes regulate cellular timekeeping and energy balance. Nature, 532, 375–379.
- Kawai N. et al. (2015). The sleep-promoting and hypothermic effects of glycine are mediated by NMDA receptors in the suprachiasmatic nucleus. Neuropsychopharmacology, 40(6), 1405–1416.
- Leproult R. et al. (1997). Sleep loss results in an elevation of cortisol levels the next evening. Sleep, 20(10), 865–870.
- Li H. et al. (2013). A genome-wide association study identifies GRK5 and RASGRP1 as type 2 diabetes loci in Chinese Hans. Diabetes, 62(1), 291–298.
- Mizuno M. et al. (2003). Association of beta-adrenoceptor polymorphisms with cardiac autonomic modulation in Japanese males. Journal of Cardiovascular Pharmacology, 42(2), 416.
- Roenneberg T. et al. (2012). Social jetlag and obesity. Current Biology, 22(10), 939–943.
- Takahashi J.S. (2017). Transcriptional architecture of the mammalian circadian clock. Nature Reviews Genetics, 18(3), 164–179.
- Xie L. et al. (2013). Sleep drives metabolite clearance from the adult brain. Science, 342(6156), 373–377.
- Brzezinski A. et al. (2005). Effects of exogenous melatonin on sleep: a meta-analysis. Sleep Medicine Reviews, 9(1), 41–50.
- Samarin S. (2026). The EscapeMed 30D System: Signal Logic, Dose Rationale, and Pilot Evidence. Escape Protocol Research, Preprint. escapeprotocol.com